Type 1 Secretion Systems
General
The secretion of proteins is an inevitable task for prokaryotic organisms to interact with their surrounding environment. Being involved in nutrient acquisition, mobility, membrane biogenesis and pathogenesis, extracellular peptides are important for many aspects of an organism’s survival and adaptation to their ecological niche. Gram-negative bacteria are equipped with at least seven dedicated secretion systems that mediate the export of proteins beyond the outer membrane. These are called Type 1 to 6 and Type 9 secretion systems (T1SS to T6SS and T9SS). The two membranes present in Gram-negative bacteria are overcome either by one-step transport mechanisms (Type I and Type III), where the unfolded substrate is translocated directly into the extracellular space without any periplasmic intermediates or by two-step mechanisms (Type II and Type V), where the substrate is first transported into the periplasm to allow folding before a second transport step across the outer membrane occurs. Additionally, T3SS, T4SS and T6SS are even capable of delivering their cargo directly into the cytosol of the host cell.
One of the most prominent members of the T1SS is the hemolysin A (HlyA) secretion system from Escherichia coli (E. coli). The substrate, the hemolytically active exotoxin HlyA, is secreted unfolded in one step, without periplasmic intermediate, across both membranes. T1SS generally comprise a relatively simple assembly of an ABC transporter and a membrane fusion protein in the inner membrane, and a TolC-like outer membrane protein. In the case of HlyA-T1SS, these are hemolysin B (HlyB), hemolysin D (HlyD) and TolC, respectively. While HlyB and HlyD form a complex in the inner membrane, TolC, which is also involved in many other export processes, is only recruited upon substrate recognition in the cytosol. Canonical ABC transporters are composed of four modules, two transmembrane domains (TMD) and two nucleotide-binding domains (NBD). As the hlyB gene encodes only one TMD and one NDB, HlyB is suggested to be functional as homodimer. Additionally, HlyB contains an N-terminal domain of 127 amino acid residues, which sequence classifies it as a member of the C39 peptidase family. The architecture of the HlyA T1SS is shown in Figure 1.
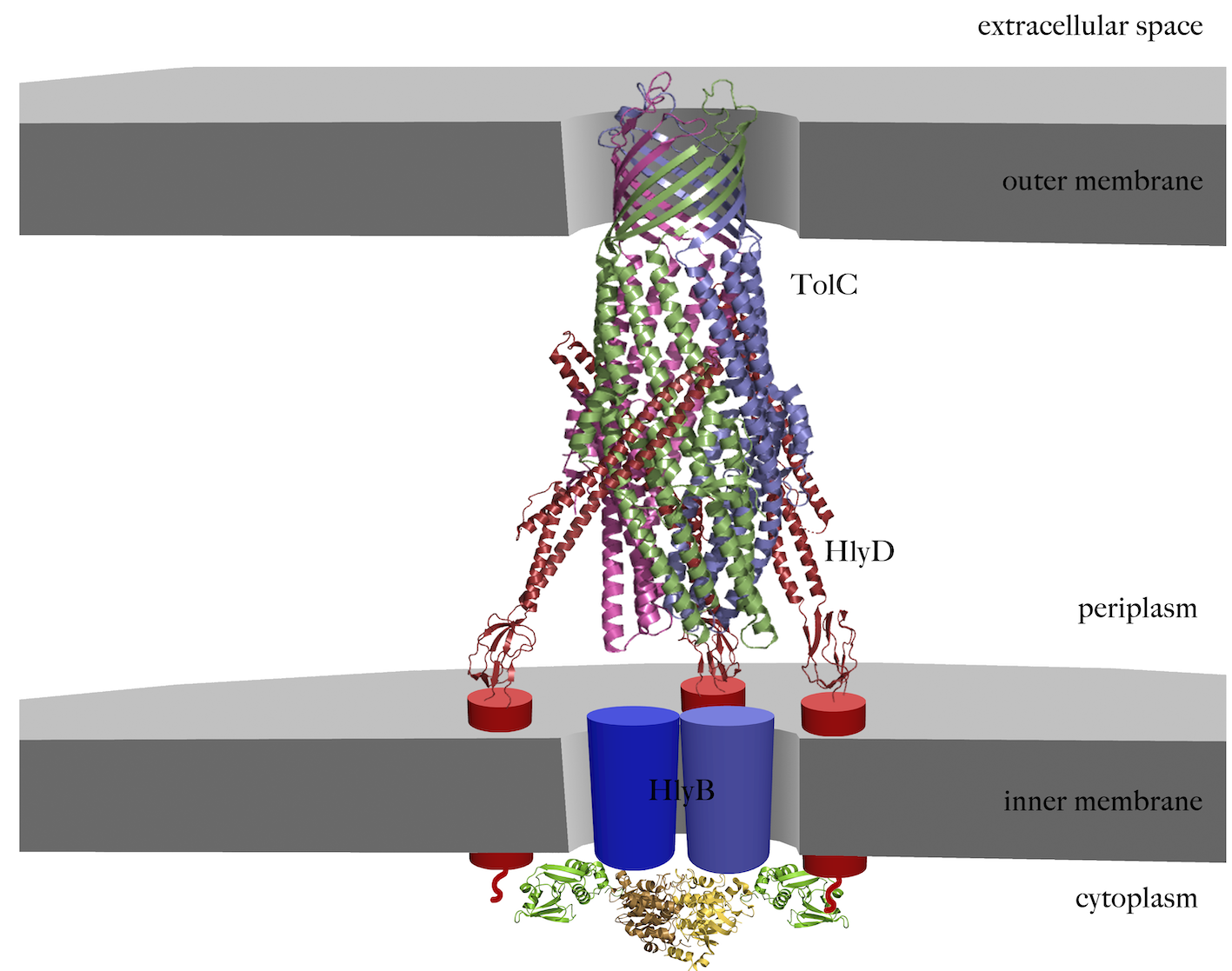
Figure 1 Schematic representation of the assembled HlyA T1SS. The trimeric crystal structure of TolC (pdb code 1ek9) in the outer membrane is shown in magenta, green and blue cartoons. The crystal structure of the known part of HlyD (pdb code 5c21) is shown in red cartoon, while symbols represents the unknowns parts, the transmembrane helix and the 60 amino acid large cytoplasmic part. The structures of the CLD (pdb code 3zua) and the NBD (pdb code 1xef) are summarized as green and yellow cartoons, respectively. The unknown TMDs is highlighted as blue cylinders,
The substrate HlyA contains a secretion signal that has been located to the last 50-60 C-terminal amino acids. A C-terminal secretion signal is generally the case in all substrates of T1SS. Additionally, HlyA belongs to the family of RTX-proteins (“repeats in toxin”), whose characteristic is the presence of a variable number of nonapeptide repeats (RTX-domains, consensus sequence of the repeat is GGXGXDXUX (X: any amino acid residue, U: large, hydrophobic residue)) that bind calcium ions and trigger folding of the protein in the extracellular space. As the cytosolic concentration of Ca2+ is too low to allow binding of Ca2+ to the nonapeptide repeats, substrates of T1SS remined unfolded in the cytosol, despite their impressive size (1024 amino acids in the case of HlyA).
Cellular studies
In T1SS, it was not known whether transport occurs directionally, i.e. the N- or the C-terminus of T1SS substrates is secreted first. To address this question we constructes HlyA fusions with the rapidly folding eGFP resulting in a stalled T1SS. Differential labeling and subsequent fluorescence microscopic detection of C- and N-terminal parts of the fusions allowed us to demonstrate vectorial transport of HlyA through the T1SS with the C-terminus appearing first outside the bacterial cells. Furthermore, we demonstrated that ATPase activity of HlyB was important in an early step of secretion as no substrate reached the cell envelop when we used an ATPase deficient HlyB mutant. This is in clear contrast to the majority of ABC transporter. Here, ATP hydrolysis is thought to reset the system.
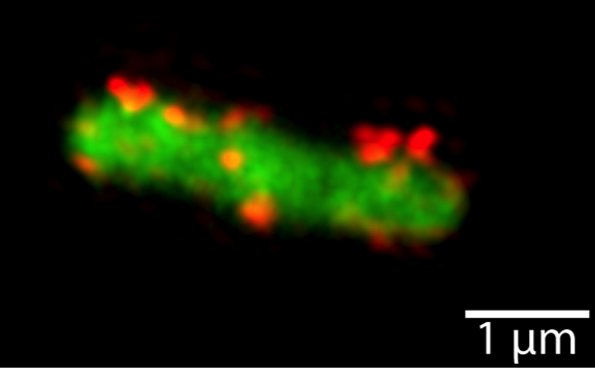
Figure 2 Detection of the surface exposed HlyA fragment of eGFP-HlyA by structured illumination microscopy (SIM). Shown are maximum intensity projections of z-stacks of a representative E. coli cell expressing eGFP-HlyA together with HlyB and HlyD. The eGFP fluorescence (green) is displayed in wide field mode, the HlyA signal (red) is displayed in high resolution mode after SIM processing.
The concept of a stalled T1SS was even extended to determine the rate of secretion per T1SS. It allowed us to determine the number of active T1SS per cell. This knowledge was used to calculate the secretion rate of the HlyA T1SS to 16 amino acids per transporter and second. It other words, it requires approximately one minute to secrete a HlyA molecule to the cell exterior and the HlyA T1SS is now one of the few bacterial secretion systems, for which quantitative data is available. For a long time the driving force for substrate secretion was discussed. Intuitively, it was always assumed that the binding of Ca2+ in the extracellular space and folding of the substrate represents this force and also prevents back sliding of the substrate during secretion. We could however demonstrate that Ca2+ has no influence on the secretion rate in the HlyA T1SS, which remained at 16 amino acids per transporter and second even in the absence of any extracellular Ca2+.
Functional studies
C39 peptidases are thiol-proteases of the papain superfamily and have been described for ABC transporters dedicated to bacteriocin export. The catalytic center of C39 peptidases contains two highly conserved residues, a cysteine and a histidine. However, sequence alignments of HlyB with various bacteriocin ABC transporters revealed that the conserved cysteine is replaced by a tyrosine residue. This suggests that the C39 peptidase of HlyB is inactive. Therefore, we named it the "C39-like domain" (CLD). We investigated the CLD of the ABC transporter HlyB structurally (see below) and functionally in great detail. We were able to show that the presence of the proteolytically inactive CLD in HlyB is essential for the secretion of HlyA and that the isolated CLD interacts specifically with unfolded HlyA, independently of the secretion signal. In combination with NMR studies (see below) we pin-pointed the substrate binding site of the CLD.
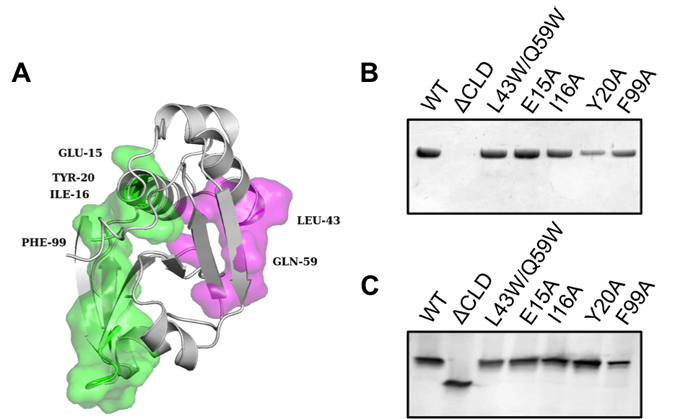
Figure 3 (A) Cartoon representation of the NMR structure of the CLD (grey). Residues forming the substrate-binding interface of ComA-PEP are shown as magenta surface. Leu43 and Gln59 of the HlyB-CLD correspond to Ala51 and Ala67 in the ComA-PEP. Amino acids of the CLD that displayed the largest changes in the CSP experiments with HlyA2 are shown as green surface. The amino acids that were mutated in this study are indicated. (B) Western blotting of supernatant samples after secretion analysis with wild-type HlyB and mutated HlyB using a polyclonal antibody raised against HlyA. The HlyB mutations are indicated. (C) Western blotting of cell lysates after the secretion analysis of (B) with a polyclonal antibody raised against HlyB.
The bottleneck in studying membrane proteins in vitro is often related to the requirement to purify membrane proteins to high purity and homogeneity. When using detergents for their extraction from membranes, membrane proteins are pulled out of their natural environment. We reconstituted HlyB into saposin-based nanoparticles. This system was suitable for the functional characterisation of the ABC transporter. Equally important, reconstitution in nanoparticles was achieved not only in the presence of lipids, but for the first time also in the presence of detergent-like lysolipids. The functional data of reconstituted HlyB revealed important differences from those obtained for detergent-solubilised HlyB, especially in comparison to a truncated mutant lacking the CLD, and highlights the impact of free detergent in buffer solution on the activity of HlyB. More important, the data revealed a modulation of the activity of HlyB by lipids in the presence of the substrate HlyA.

Figure 4 (A) Silver-stained SDS PAGE gel of HlyB reconstituted in DOPC/saposin nanoparticles (left panel) and DOPC/saposin (right panel). Saposin is abbreviated Sap-A. (B) Unfolded pro-HlyA mediated modulation of ATPase activity of HlyB reconstituted in DOPC/saposin (black squares) and LPC/saposin (blue circles).
Structural studies
The structure of the CLD was solved by NMR spectroscopy, which allowed a detailed comparison with catalytically active C39 peptidases and the mapping of the ligand-binding site by chemical shift perturbation (CSP) experiments. Taken together, our results provide the first insights into the function of a CLD and highlight its essential role in the initial recognition events of a T1SS. We propose that the CLD shields HlyA to prevent the aggregation and/or degradation of the secretion-competent, unfolded substrate.
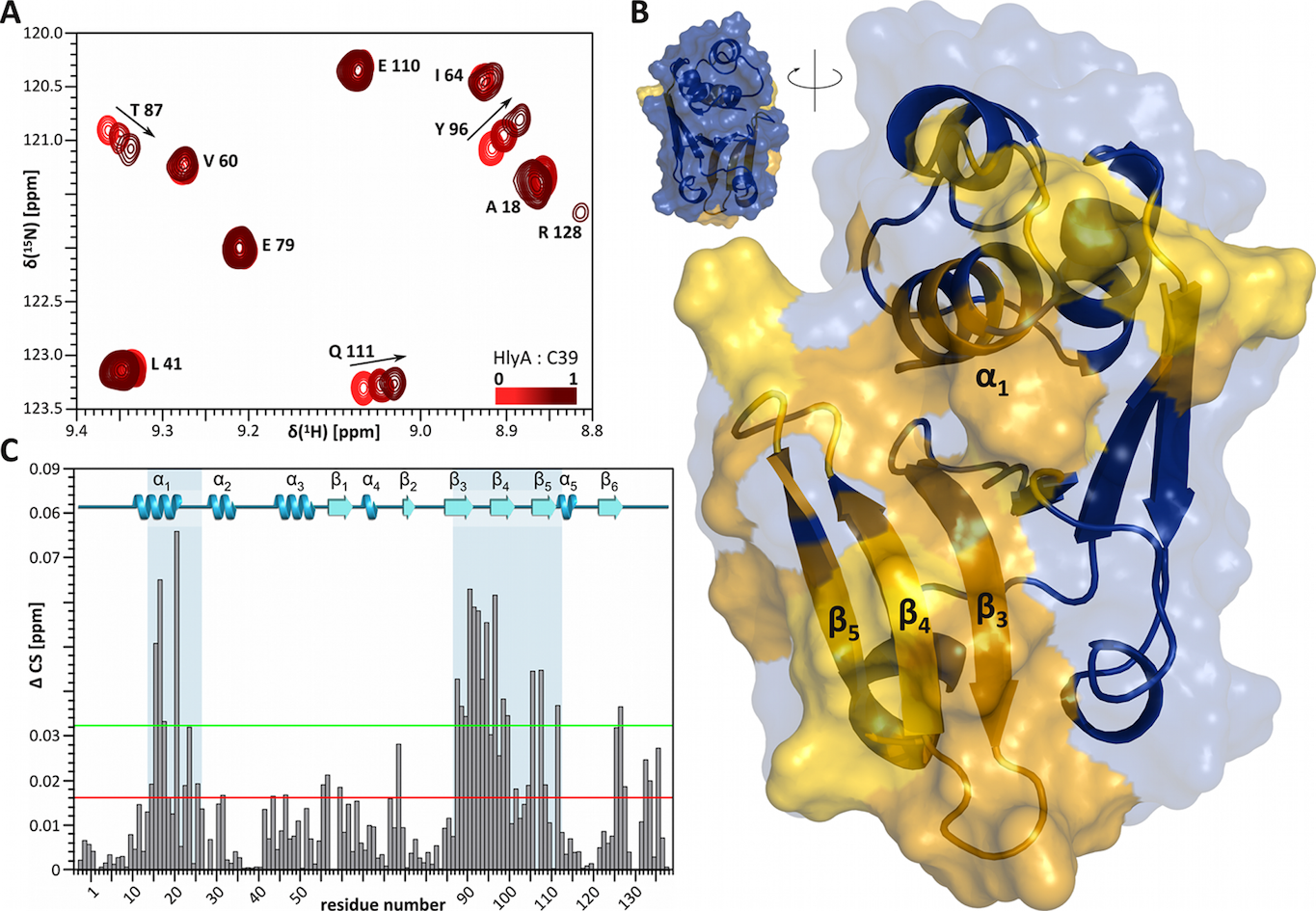
Figure 5 Interaction of HlyA with the CLD characterized by NMR chemical shift perturbation (CSP) experiments. (A) Part of the HSQC spectra illustrating the CSP. Starting with an equimolar mixture of HlyA and CLD (75 µM), the CLD concentration was kept constant, while HlyA was diluted in a three step series, each step reducing the HlyA concentration to a third. Arrows highlight the trajectory of chemical shift changes. (B) The chemical shift changes were mapped onto the CLD structure. Interacting regions are highlighted in yellow, while non-interacting regions are shown in blue. (C) The chemical shift changes were mapped onto the primary sequence of the CLD.
In addition to the NMR studies, we have performed detailed structural and biochemical studies on the isolated NBD of HlyB, which are summarized in Figure 6.
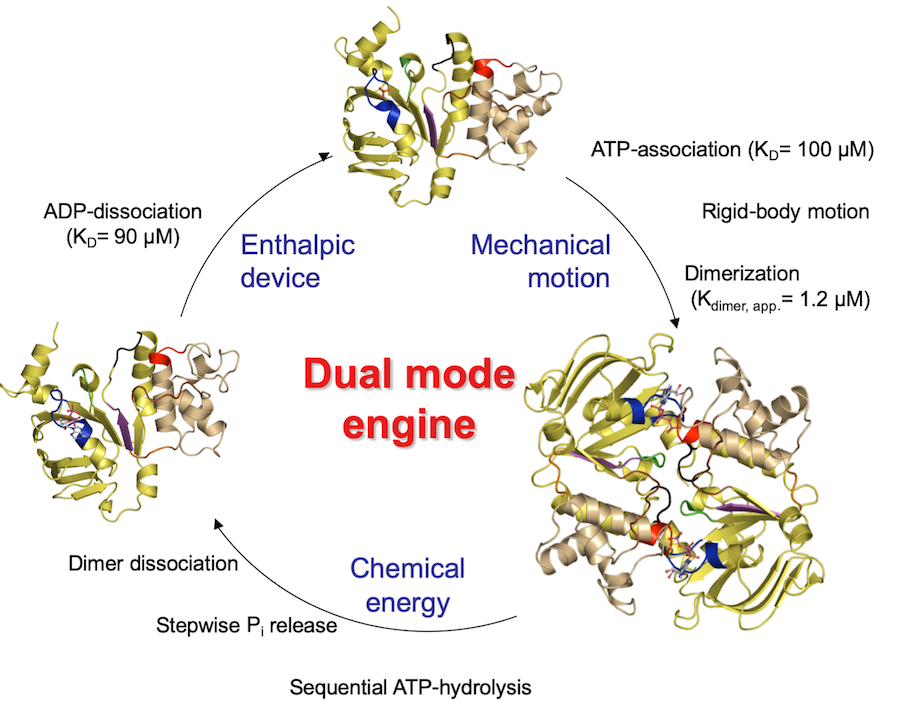
Figure 6 The catalytic cycle of the isolated NBD of HlyB.
Biotechnological applications
In addition we try to use the HlyA T1SS as platform for the biotechnological production of proteins. A biotechnological production of proteins through protein secretion systems is often superior to the conventional cytoplasmic production, because of the absence of large amounts of proteases present in the extracellular space and the ease of purification or downstream processing. Recently, we revealed that the folding rate of the passenger protein dictates secretion efficiency in T1SS. Here, the well-known MalE or IFABP (intestinal fatty acid binding protein) failed to be secreted when fused to a C-terminal fragment of the natural substrate HlyA. In contrast, slow-folding mutants of MalE or IFABP were secreted in high yields and, importantly, in a biological active state.
In continuation of our interest to exploit and optimize the HlyA T1SS for the production of heterologous proteins, we identified an A/U rich enhancer region upstream of hlyA, required for effective expression and secretion of selected heterologous proteins irrespective of their prokaryotic, viral or eukaryotic origin. We further demonstrated that ribosomal protein S1 binds to the hlyA A/U rich enhancer region and that this region was involved in the high yields of secretion of functional proteins. This concept has now been applied to a large variety of proteins resulting of successful secretion of most of the investigated systems.

Figure 7 Schematic representation of the hly operon and the effect of the enhancer region on the expression of HlyA1. (a) The hly operon consists of four genes in the order - hlyC, hlyA, hlyB and hlyD. Downstream of the promoter (depicted with a black arrow), a JUMPstart (shown in grey box) and an ops sequence (represented by a black box) are present. The stem loop termination sequence located between the hlyA and hlyB genes is shown as a kink. (b) The AU-rich upstream region of hlyA is highlighted in the box. The start codon of hlyA (AUG) and the stop codon of hlyC (UAA) are shown in bold while the ribosome-binding site is underlined. (c) SDS-PAGE analysis of non-concentrated supernatant obtained after expression of hlyA1 gene containing either 159 (lane 1) or 17 (lane 2) nucleotides upstream of hlyA region and a molecular weight marker (lane M).
Selected references
Kanonenberg, K., S.H.J. Smits & L. Schmitt, (2019) Functional Reconstitution of HlyB, a Type I Secretion ABC Transporter, in Saposin-A Nanoparticles. Sci Rep 9: 8436.
Khosa, S., R. Scholz, C. Schwarz, M. Trilling, H. Hengel, K.E. Jaeger, S.H.J. Smits & L. Schmitt, (2018) An A/U-Rich Enhancer Region Is Required for High-Level Protein Secretion through the HlyA Type I Secretion System. Appl Environ Microbiol 84: e01163- 01117.
Lenders, M.H., T. Beer, S.H. Smits & L. Schmitt, (2016) In vivo quantification of the secretion rates of the hemolysin A Type I secretion system. Sci Rep 6: 33275.
Holland, I.B., S. Peherstorfer, K. Kanonenberg, M. Lenders, S. Reimann & L. Schmitt, (2016) Type I Protein Secretion-Deceptively Simple yet with a Wide Range of Mechanistic Variability across the Family. EcoSal Plus 7.
Lenders, M.H., S. Weidtkamp-Peters, D. Kleinschrodt, K.E. Jaeger, S.H. Smits & L. Schmitt, (2015) Directionality of substrate translocation of the hemolysin A Type I secretion system. Sci Rep 5: 12470.
Thomas, S., I.B. Holland & L. Schmitt, (2014) The Type 1 secretion pathway - The hemolysin system and beyond. Biochim Biophys Acta: 1621- 1649.
Lecher, J., C.K. Schwarz, M. Stoldt, S.H. Smits, D. Willbold & L. Schmitt, (2012) An RTX transporter tethers its unfolded substrate during secretion via a unique N-terminal domain. Structure 20: 1778-1787.
Bakkes, P.J., S. Jenewein, S.H. Smits, I.B. Holland & L. Schmitt, (2010) The rate of folding dictates substrate secretion by the Escherichia coli hemolysin type 1 secretion system. The Journal of biological chemistry 285: 40573-40580.
Zaitseva, J., C. Oswald, T. Jumpertz, S. Jenewein, A. Wiedenmann, I.B. Holland & L. Schmitt, (2006) A structural analysis of asymmetry required for catalytic activity of an ABC-ATPase domain dimer. Embo J 25: 3432-3443.
Zaitseva, J., S. Jenewein, T. Jumpertz, I.B. Holland & L. Schmitt, (2005) H662 is the linchpin of ATP hydrolysis in the nucleotide-binding domain of the ABC transporter HlyB. Embo J 24: 1901-1910.